CAPITULO 4CIRCULACION PULMONAR |
|
|
Presiones en el circuito menorLas presiones de la aurícula y ventrículo derechos y de la arteria pulmonar pueden medirse directamente mediante un catéter introducido hasta estas cavidades a través de una vena de las extremidades o del cuello. De uso más frecuente en clínica es el catéter de Swan Ganz, que está provisto de un pequeño balón inflable en su extremo distal, el cual flota y es arrastrado por la corriente sanguínea hasta alcanzar la arteria pulmonar, lo que permite su introducción sin necesidad de control radioscópico. Si el catéter se avanza hasta ocluir una rama arterial, la presión que se registra, llamada de enclavamiento o de capilar pulmonar, corresponde prácticamente a la de la aurícula izquierda. Este catéter tiene frecuente aplicación en las unidades de tratamiento intensivo en pacientes en los cuales se necesita monitorizar las presiones del circuito menor para fines diagnósticos o terapéuticos. En clínica también puede obtenerse una medición suficientemente aproximada con la ecografía cardiaca Las presiones en el circuito pulmonar son aproximadamente seis veces menores que las del circuito sistémico: la presión media en la aorta es de 100 mmHg, mientras que en la arteria pulmonar es de sólo 15 mmHg. En concordancia, las paredes de las arterias pulmonares son muy delgadas y están provistas de muy escasa musculatura lisa, confundiéndose fácilmente con venas de diámetro similar. Esta característica anatómica contrasta con las arterias sistémicas, que tienen paredes gruesas y abundante musculatura lisa, especialmente a nivel arteriolar. Si se tiene presente las diferentes funciones de estas dos circulaciones, se explica que sus regímenes de presiones sean diferentes. La circulación sistémica suministra sangre a todos los órganos, incluso cuando están ubicados por sobre el nivel del corazón, como por ejemplo la cabeza o un brazo elevado, y redistribuye por vasoconstricción selectiva la sangre de una región a otra de acuerdo a los requerimientos metabólicos de los tejidos. En cambio, en el pulmón la presión arterial sólo necesita alcanzar el nivel necesario para impulsar la sangre hasta los vértices, que en posición de pie están sólo a 15-20 cm por encima del tronco de la arteria pulmonar. Resistencia vascular pulmonarEl flujo de sangre a través del circuito pulmonar está determinado por la diferencia de presiones entre el tronco de la arteria pulmonar y la aurícula izquierda. A este flujo se opone una resistencia vascular de tipo friccional, que depende de la longitud del vaso (l), la viscosidad de la sangre (µ) y del radio (r) (ecuación de Poiseuille).
En esta fórmula es evidente que el factor más crítico para determinar la resistencia vascular pulmonar (RVP) es el calibre del vaso, debido a que la resistencia es inversamente proporcional a su radio elevado a la cuarta potencia. De esta forma, una disminución de un 50% en el radio de un vaso, como puede ocurrir por vasoconstricción, aumenta su resistencia 16 veces. Esta fórmula se aplica en forma rigurosa cuando se trata de tubos rígidos perfundidos por un líquido homogéneo que fluye en forma laminar y continua, pero los vasos sanguíneos son distensibles, están perfundidos por sangre que no es homogénea y con un flujo pulsátil, de manera que su aplicación en fisiología y clínica permite sólo resultados aproximados. La RVP puede también calcularse de acuerdo a la ley de Ohm, como se hace corrientemente en la práctica clínica. De acuerdo a esta ley la resistencia está determinada por la siguiente ecuación:
(15 - 8) / 5 = 1,4 mmHg/L/min
En estudios hemodinámicos la presión se mide en dinas/cm2(fuerza por unidad de área) y el flujo sanguíneo se determina en cm3/segundo. En el hombre normal la resistencia vascular pulmonar fluctúa entre 100 a 200 dinas x s/cm5. En condiciones normales, las arterias pulmonares de pequeño calibre y los capilares son los vasos que ofrecen la mayor resistencia al flujo sanguíneo, a diferencia de la circulación sistémica, donde la mayor resistencia se encuentra al nivel de las arteriolas. El sistema venoso pulmonar ofrece escasa resistencia, con un mínimo gradiente de presión entre capilar pulmonar y aurícula izquierda. Si bien la resistencia vascular pulmonar en reposo es baja, ella puede disminuir aun más si aumenta el flujo en los vasos pulmonares, como ocurre durante el ejercicio. Dos mecanismos son responsables de este efecto: A medida que la presión sube, comienzan a abrirse capilares que, en reposo, estaban cerrados o con muy escaso flujo, es decir, se reclutan nuevos vasos bajando la resistencia (Figura 4-1). 
La razón de que sólo una parte de los vasos están abiertos en reposo no es clara, pero puede deberse a características geométricas de la red que forman, existiendo canales preferenciales de flujo. Otra posible explicación es que exista una presión crítica de apertura, que debe ser sobrepasada para que la sangre pueda fluir. El segundo mecanismo es la distensión de los vasos, con el consiguiente aumento de su calibre, fenómeno que es el principal responsable de la caída de resistencia. Estos mismos explicarían que la presión de la arteria cambie escasamente después de una neumonectomía que reduce el lecho vascular pulmonar a la mitad. Durante el ejercicio, estos mecanismos de reclutamiento
y distensión son los que permiten acomodar, prácticamente
sin cambios en la presión, el aumento del gasto cardíaco
y del volumen sanguíneo pulmonar.
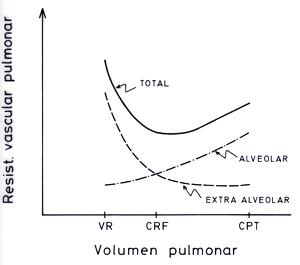
Flujo sanguíneo pulmonarLa circulación pulmonar recibe la totalidad del volumen sistólico del ventrículo derecho, que en condiciones normales es prácticamente igual al del ventrículo izquierdo. En la práctica clínica, el flujo sanguíneo pulmonar se mide mediante termodilución. Para ello se usan catéteres de tipo Swan-Ganz, que están equipados con un pequeño sensor de temperatura en su extremo distal y con un lumen adicional, a través del cual se inyecta suero helado. Al inyectar el suero, el sensor detecta una disminución de la temperatura, cuya duración e intensidad depende del flujo sanguíneo. Conociendo la temperatura del bolo inyectado, su volumen y el área bajo la curva temperatura-tiempo, se puede calcular el gasto cardíaco. En el adulto normal, el gasto cardíaco, y por lo tanto el flujo sanguíneo pulmonar, fluctúa entre 5 a 8 L/min. Si el gasto cardíaco se relaciona con la superficie corporal se obtiene el índice cardíaco, cuyos valores varían entre 2,7 y 3,2 L/min/2. Características funcionales de los vasos sanguíneos pulmonaresEl grado de dilataciónde cualquier vaso es determinado por su presión transmural, que se define como la diferencia de presión que actúa a través de su pared, esto es, la presión intraluminal (Pi) menos la presión perivascular o externa (Pe). Debido a la gran distensibilidad de las arterias pulmonares el rango de variación de su diámetro es muy amplio Las presiones peri vasculares dependen de las estructuras que rodean al vaso. Los grandes vasos extrapulmonares están sujetos a las fluctuaciones de la presión pleural. Los vasos intrapulmonares están sujetos a diferentes regímenes de presiones dependiendo de su localización alveolar o extra alveolar. Los vasos extraalveolares son afectados simultáneamente por la presión pleural y la presión del intersticio. Los vasos alveolares o capilares ubicados en los septa interalveolares están sujetos a los regímenes de presiones de los alvéolos que los rodean, sin que los afecten los cambios de la presión pleural. Factores mecánicos que regulan la resistencia vascular pulmonarPor sus características estructurales, su ubicación dentro del tórax e interposición entre ambos ventrículos, la circulación pulmonar normal está sometida a múltiples influencias extravasculares, a las que responde pasivamente con gran sensibilidad. Por ello, en su regulación predominan los efectos de tipo mecánico sobre aquellos propios de una actividad vasomotora propiamente tal. Presiones transmurales. Un aumento de Pi (como se observa cuando aumenta la presión de aurícula izquierda) aumenta la presión transmural y dilata los vasos pulmonares, reduciendo la RVP. Volumen pulmonar. Al inspirar desde volumen residual a capacidad pulmonar total, la resistencia de los vasos alveolares aumenta progresivamente, mientras que lo contrario ocurre con los vasos extra alveolares. Esto se debe a que los vasos alveolares son comprimidos y elongados a medida que el pulmón se insufla, mientras que la presión intersticial que rodea a los vasos extra alveolares se hace más negativa, causando una disminución de Pe y por lo tanto un aumento de su presión transmural. Lo contrario ocurre al nivel de volumen residual, condición en que los vasos alveolares se encuentran dilatados y los vasos extra alveolares, debido al aumento de la presión pleural, colapsados (Figura 4-2).
Durante la respiración tranquila los vasos extra alveolares están expuestos siempre a una presión externa o perivascular que es más negativa que la de los vasos alveolares, lo que tiene importancia en la formación del edema pulmonar, que comienza siempre en el territorio extra-alveolar por sus mayores presiones transmurales. Esto determina que en las etapas iniciales de la formación de edema la hematosis no se ve afectada, como veremos en el capítulo correspondiente. Efecto de la fuerza de gravedad.
El bajo régimen de presiones que existe en la circulación
pulmonar la hace muy sensible a la influencia de la gravedad,
fenómeno que no se observa en la circulación
sistémica. La consiguiente falta de uniformidad en
la perfusión del pulmón fue analizada en el
capítulo sobre relación Viscosidad sanguínea.
La viscosidad sanguínea depende del hematocrito, de
la deformabilidad de los eritrocitos y de la composición
del plasma. Si bien es difícil demostrar la magnitud
con que cada uno de estos factores influye sobre la resistencia,
existen evidencias que demuestran que el aumento de viscosidad
producido por el aumento de eritrocitos es un factor importante
de aumento de la resistencia vascular pulmonar. Factores vasomotores que regulan la resistencia vascular pulmonarEstímulos neurogénicos. Pese a que las arterias y venas pulmonares están inervadas con fibras de origen simpático y parasimpático, esta inervación es claramente menor que la de la circulación sistémica. Estudios experimentales y clínicos han llegado a la conclusión de que el sistema nervioso autónomo no intervendría en forma notoria en el control de la circulación pulmonar en el adulto normal. Estímulos humorales. Numerosos vasoconstrictores y vasodilatadores pulmonares han sido identificados en los años recientes. Sin embargo, en la gran mayoría de los casos su conocimiento sólo tiene interés para el investigador. Desde un punto de vista clínico, es importante destacar que las catecolaminas epinefrina, norepinefrina, dopamina utilizadas corrientemente en las unidades de cuidados intensivos producen vasoconstricción pulmonar. Como vasodilatadores se recurre al óxido nítrico y a drogas de empleo habitual, como la dobutamina y el isoproterenol, entre las catecolaminas, y a nitroprusiato, nitroglicerina, hidralazina, prostaglandinas y sildenafil entre los vasodilatadores directos. Estímulos bioquímicos.
La hipoxia alveolar es el más importante vasoconstrictor
de las arteriolas pulmonares, a través de un mecanismo
local (vasoconstricción pulmonar hipóxica).
En condiciones normales, este fenómeno es capaz de
corregir en parte los trastornos
BALANCE HIDRICO PULMONARLa mantención de una determinada cantidad de liquido en el intersticio pulmonar y dentro de los alvéolos es de suma importancia ya que su acumulación excesiva rigidiza el pulmón e interfiere con la hematosis con graves consecuencias. Los mecanismos que controlan este aspecto serán resumidos a continuación. Filtración transcapilarLa filtración de líquidos a través de la pared capilar se describe mediante la ecuación de Starling: Flujo transcapilar = Kfc [(Pc - Pi)
- El término Kfc corresponde al coeficiente
de filtración de la membrana capilar dependiente de
su conductividad y su área; Pc y Pi son respectivamente
las presiones hidrostáticas capilar e intersticial,
mientras que πc y πi
se refieren a las presiones oncóticas capilar e intersticial,
respectivamente. El símbolo La medición de todas estas presiones es difícil, incluso en condiciones experimentales y los cálculos indican que , la presión neta de filtración es de aproximadamente +8,5 mmHg, lo que es mayor que la presión de absorción, por lo que existe un flujo constante de líquido desde el espacio intravascular al intersticio. Gran parte de este líquido es removido por el sistema linfático pulmonar, que lo drena a la vena cava, pudiendo alcanzar flujos de hasta 50 ml/h. Si aumenta la permeabilidad capilar, acercándose
. |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |