|
|
|
| |
Si bien en clínica es infrecuente que
se necesite medir individualmente los mecanismos que se
analizaran en este capítulo, los conceptos involucrados
son esenciales para comprender y manejar múltiples situaciones.
Para mantener niveles normales de PaO2
y PaCO2
en las variadas condiciones de demanda metabólica, la ventilación
alveolar es regulada por una red de centros y vías neuronales
que genera los estímulos que activan rítmicamente,
no sólo los músculos respiratorios , sino también
los músculos faríngeos que mantienen abierta la
vía aérea superior. Aparte de esta función
específica, el control de la ventilación opera también
en otras actividades de naturaleza muy diferente, como son la
fonación, la risa, el llanto, la tos, los estornudos, la
regulación del pH, etc.
En este capítulo nos centraremos en aquellos
aspectos relacionados con la homeostasis gaseosa que tienen una
proyección sobre los problemas clínicos, sin entrar
en detalles morfológicos y funcionales del sistema regulador,
que pueden ser consultados en los textos especializados.
Para analizar esta función consideraremos
las organizaciones neuronales que elaboran e integran los estímulos,
las vías que los conducen y los mecanismos aferentes que
informan sobre las demandas metabólicas y retroalimentan
el sistema.
CENTROS DE LA RESPIRACION
La alternancia de inspiración y espiración
es determinada por actividad rítmica y automática
de centros nerviosos situados en el tronco cerebral. Estos centros
reciben información de otras zonas del cerebro y de sensores
situados en diversas partes de organismo , todo lo cual permite
que la respiración se adecúe a variadas demandas
metabólicas y se coordine para realizar otras funciones
diferentes, como la fonación, deglución, risa,
tos. etc.
La inspiración comienza por activación
de las neuronas inspiratorias, que envían impulsos a
la musculatura inspiratoria, básicamente el diafragma,
alrededor de cada 5 segundos en el adulto.
Los músculos faríngeos, que estabilizan
y mantienen abierta la vía aérea superior, se
activan antes que el diafragma y los intercostales, lo que impide
que la faringe se colapse por efecto de la presión negativa
generada durante la inspiración , mecanismo que puede
alterarse patológicamente durante el sueño.
Durante la respiración normal en reposo, la musculatura
espiratoria casi no participa, ya que la espiración es
un evento pasivo debido a la retracción elástica
del pulmón distendido por la inspiración previa.
Durante la primera parte de la espiración, persiste alguna
activación de los músculos inspiratorios que aminoran
la velocidad del flujo aéreo en la primera parte de la
espiración, la cual cesa durante el resto de la espiración.
Los músculos espiratorios son estimulados sólo
cuando aumenta la demanda ventilatoria en forma importante.
MECANISMOS AFERENTES
La información moduladora llega al sistema
neuronal por las vías humoral y nerviosa. La sensibilidad
de estos mecanismos varía entre individuos y parece estar
genéticamente determinada. También se producen
adaptaciones adquiridas, como las que ocurren con el crecimiento,
con el ejercicio intenso habitual o con la residencia en la
altura.
REGULACION HUMORAL
Los principales moduladores humorales de
la ventilación son los niveles arteriales de O2,
CO2 e hidrogeniones.
CONCENTRACION DE CO2 E
HIDROGENIONES
La capacidad de responder a niveles anormales
de PaCO2 con cambios en la
ventilación alveolar es fundamental para la homeostasis
ácido-base y, por lo tanto, la función celular.
y es un mecanismo muy sensible: una alza de PaCO2
de 40 a 50 mmHg sube la ventilación a 30 litros/min.
La hipercapnia aumenta tanto la actividad
de las motoneuronas que controlan los músculos de la
bomba respiratoria, como de las que estimulan de los músculos
faríngeos, que abren la vía aérea.
Para evaluar, en parte, este aspecto se usa
la medición de la respuesta ventilatoria a la inhalación
de CO2 . La inhalación
de concentraciones crecientes de CO2
produce un aumento de la ventilación, tanto
por aumento del volumen corriente como de la frecuencia respiratoria
(Fig. 7-1).
Figura 7-1. Respuesta ventilatoria
al CO2. Los sujetos inspiran mezclas gaseosas con concentraciones
elevadas de CO2. La ventilación (  E E)
aumenta en forma lineal dentro del rango estudiado y la
respuesta se evalúa mediante la pendiente de la
recta ( E/PCO 2).
Esta respuesta se potencia en presencia de hipoxemia.
La respuesta es variable de un individuo a otro y se mide
a través del cambio de ventilación en litros
por minuto, provocado por cada mmHg de aumento de la PaCO2.
El promedio normal es de 2.5 L/min/mmHg.
Existen dos tipos de receptores para el CO2:
centrales y periféricos.
a) centrales:
son los más sensibles al CO2
y sus células receptoras están principalmente
ubicadas en el bulbo, en contacto con el líquido cefalorraquídeo
(LCR). Sus neuronas son diferentes de las inspiratorias y
espiratorias, a las cuales envían su información.
Para algunos investigadores el estímulo específico
sería el CO2 mismo;
para otros sería el cambio de concentración
de hidrogeniones que el CO2
determina en el LCR.
b) periféricos:
Aunque son básicamente sensores de hipoxemia, también
responden a los cambios de pH y PaCO2
Están ubicados principalmente en los cuerpos
carotídeos y, en menor cantidad, en los cuerpos aórticos.
Su información es transmitida a los centros respiratorios
a través del noveno y décimo pares craneanos.
.Como la mayor parte de la respuesta ventilatoria al CO2
se mantiene luego de la denervación de los quimiorreceptores
periféricos, se deduce que los receptores centrales
al CO2 son los más
importantes en esta respuesta.
Por la existencia de una barrera hematoencefálica
de permeabilidad selectiva, el equilibrio ácido-base
en el LCR experimenta un desfase con el de la sangre. Entre
los factores que influyen en este fenómeno están:
-
• El poder tapón del LCR
es bajo, por su escasa cantidad de proteínas
y por la ausencia de hemoglobina.
- Los iones HCO3-
difunden muy lentamente desde la sangre al LCR, por lo que
en las acidosis y alcalosis metabólicas el pH del
LCR inicialmente no varía o lo hace, incluso, en
sentido contrario al de la sangre.
- El CO2,
en cambio, difunde libremente, modificando en forma rápida
el pH del LCR. Así, un aumento de PaCO2
lleva a un aumento paralelo de la PaCO2
en el LCR, lo que produce un cambio de pH mayor que en la
sangre por la escasez de tapones en este compartimiento.
Con el tiempo, el bicarbonato del plasma, que traspasa la
barrera lentamente y llega a corregir el pH del LCR en forma
aun más completa que en la sangre.
La disociación que se observa entre
sangre y LCR explica, como lo veremos en patología,
que la respuesta de la ventilación ante modificaciones
humorales agudas sea diferente a la que se observa en etapas
crónicas, en las que hay tiempo para que se produzcan
las modificaciones en el LCR. Estos mismos factores explican
la evolución de los fenómenos de adaptación
ventilatoria que se observan en la altura y en la insuficiencia
respiratoria crónica.
NIVEL DE OXIGENACION ARTERIAL
Al contrario de lo que ocurre con el CO2,
el nivel de oxígeno arterial es regulado por los receptores
periféricos, prácticamente sin participación
de los receptores centrales.
Quimiorreceptores periféricos
Los principales receptores periféricos
son los carotídeos, situados en la bifurcación
de la carótida. Su irrigación proviene de una
arteriola que nace directamente de la carótida y provee
a las células receptoras de un alto flujo sanguíneo
que, por gramo de tejido, es 40 veces mayor que el del cerebro.
Los estímulos específicos para su activación
son:
a) La disminución de la PaO2.
El descenso del contenido de O2
no estimula estos receptores, lo que explica la escasa respuesta
ventilatoria que se observa en reposo en la anemia y en la
intoxicación por CO, situaciones en las cuales está
disminuido el contenido de O2
pero la PaO2 es
normal.
b) La disminución del flujo
sanguíneo. Este factor actuaría a través
de los cambios locales de pH que se producen debido a una
menor remoción de metabolitos. Este mecanismo explicaría
la hiperventilación que se observa en el shock.
Los quimiorreceptores no modifican su frecuencia
de descarga hasta que la PaO2 baja hasta 50-70
mmHg, y su efecto máximo es relativamente escaso, pues
aumentan la ventilación alveolar solo en un 68% (Figura
7-2).
 |
Figura 7-2. Respuesta ventilatoria a la hipoxemia.
Los sujetos inspiran mezclas gaseosas con concentraciones
decrecientes de O2 , lo que disminuye la PaO2
. La ventilación aumenta en forma alinear
y entre 100 y 60 mmHg los cambios de PaO2
prácticamente no modifican la E.
Bajo esta cifra se produce un ascenso de la E
, primero moderado que se torna exponencial por debajo
de 40 mmHg
|
Estos receptores periféricos tienen,
en cambio, la importante característica de que no se
inhiben frente a una serie de condiciones que deprimen los
receptores bulbares, como son la propia hipoxemia, la hipercarbia,
los hipnóticos, anestésicos, etc. En estas circunstancias
la respiración del paciente pasa a depender exclusivamente
de la actividad del receptor carotídeo. Por estas características
de los quimiorreceptores periféricos, la PaO2
no es el principal factor de regulación durante
la respiración normal, pero puede llegar a ser ser
vital en condiciones en que hay depresión del sistema
nervioso central. Debe destacarse que en algunas condiciones
de hipoxemia crónica el cuerpo carotídeo puede
llegar a hipertrofiarse hasta 8 veces el tamaño normal.
La acción del O2
sobre los quimiorreceptores periféricos se potencia
si aumenta la PaCO2
o aumenta la concentración de H+.
Es interesante lo que ocurre con la respuesta
ventilatoria de un sujeto que reside a nivel del mar y que
sube a zonas de gran altitud. La ventilación aumenta
en respuesta a la disminución de la PaO2,
aunque no todo lo que cabe esperar, debido al efecto inhibitorio
que tiene la alcalosis respiratoria secundaria a la hipocapnia
causada por la misma hiperventilación. Con la aclimatación
a la altura, el organismo se adapta a una menor PaCO2
y la alcalosis se compensa por disminución del bicarbonato,
con lo que disminuyen los factores frenadores de la ventilación
y ésta puede aumentar de acuerdo a la magnitud de la
hipoxemia.
REGULACION NEUROLOGICA
CONTROL CENTRAL VOLUNTARIO
La actividad respiratoria puede ser modificada
en forma voluntaria por actividad de la corteza cerebral,
pero sólo en forma temporal, ya que los cambios fisiológicos
secundarios a la modificación ventilatoria terminan
por imponer el control automático. Una hiperventilación
voluntaria termina por mareos y pérdida de conciencia
secundarios a la alcalosis respiratoria aguda que provoca
y una apnea voluntaria tiene una duración limitada
por la hipercarbia
CONTROL CENTRAL INCONSCIENTE
. Muchos actos automáticos, como la
deglución, fonación, tos, preparación
para el ejercicio, etc., involucran una adecuación
automática del ritmo respiratorio que es comandada
por vía neurológica. Dentro de esta misma categoría
cae el jadeo utilizado para el control del calor por algunos
animales de pelo largo o que carecen de glándulas sudoríparas,
en los cuales el aumento de ventilación superficial
sirve para disipar calor sin que se produzca alcalosis respiratoria,
porque se ventila sólo el espacio muerto. Además,
es una observación común que las emociones influyen
marcadamente en la ventilación
REFLEJOS DE ORIGEN PULMONAR
Existen tres tipos de receptores en el pulmón,
cuya información va a los centros respiratorios a través
del vago:
Receptores de distensión. Son
de adaptación lenta y se encuentran en relación
con el músculo liso de la vía aérea,
cuya elongación en inspiración sería
el estímulo específico. Uno de los efectos de
la vía refleja en la que participan es la frenación
de la inspiración (reflejo de Hering-Breuer) que en
algunos animales regularía la alternancia de las fases
respiratorias, pero que en el hombre es activo sólo
en el recién nacido o cuando se respira con volúmenes
corrientes muy grandes.
Receptores de irritación.
Son de adaptación rápida y se les supone
una finalidad primariamente defensiva. Son estimulados por
gases irritantes, estímulos mecánicos, histamina,
reacciones alérgicas, congestión pulmonar pasiva,
embolia pulmonar, etc. Su respuesta es la broncoconstricción,
constricción laríngea y tos. Se localizan preferentemente
en la laringe y vías aéreas centrales, aparentemente
en relación con el epitelio.
Receptores J o juxtacapilares. Son
receptores localizados en el intersticio alveolar, en la cercanía
de los capilares. Se estimulan por procesos que comprometen
esta área, tales como el edema intersticial y la acción
de irritantes químicos. Contribuirían a la taquipnea
y a la sensación de disnea que acompaña a estas
condiciones. Los impulsos de los receptores de irritación
son enviados al centro respiratorio a través del vago.
RECEPTORES MUSCULARES
Los músculos intercostales y el diafragma
poseen husos musculares que captan el grado de elongación
del músculo. Esta información es importante
para el control reflejo de la fuerza de contracción.
Se ha planteado que estos receptores intervienen en la sensación
de disnea cuando captan que el esfuerzo muscular es excesivo
para el efecto ventilatorio que se consigue (capítulo
18). .En los músculos en ejercicio se activarían
sensores que captan condiciones locales como acidosis o falta
de aporte de oxigeno.Esto último explicaría
por que el ejercicio provoca hiperpnea en anemia, a pesar
que no lo hace en reposo.
INTEGRACION DE LOS MECANISMOS REGULADORES
En un organismo que no está sometido
a las condiciones controladas de un experimento, los mecanismos
que hemos descrito separadamente operan como un todo. Así,
la respuesta global de un individuo ante uno o más estímulos
será la suma de:
1.-La acción del o los estímulos
sobre los diferentes receptores, cuyas respuestas pueden ser concordantes
o contrapuestas.
2.-Las reacciones secundarias del organismo ante
estas respuestas.
3.-La asociación de otros estímulos
sinérgicos o antagónicos.
4.-La sensibilidad de los receptores, que puede variar por
factores concomitantes o
fenómenos de adaptación:
5.-El efecto del factor tiempo.
6.-El estado de la conducción nerviosa.
7.-El estado de los efectores de la ventilación.
Es importante tener presente esta complejidad
cuando se pretende intervenir terapéuticamente sobre
alguno de los eslabones de la cadena o cuando se quiere interpretar
un hecho patológico.
REGULACION DE LA RESPIRACION EN EL EJERCICIO
El ejercicio físico es la condición
fisiológica que con mayor frecuencia e intensidad genera
una demanda metabólica sobre el sistema respiratorio
. Para ello los centros reguladores se coordinan en una compleja
secuencia . diferente a la del reposo en que también
participan los sistemas circulatorio y nervioso
autónomo, cuyo objetivo no es reaccionar ante la alteración
de gases ya producida, sino mantener su normalidad aunque aumente
su consumo o su producción. La eficacia de este proceso
puede apreciarse en el hecho que en un ejercicio extremo el
consumo de oxigeno puede decuplicar los 250 ml/min del
reposo.
Aunque no todas sus fases y mecanismos tienen
un comprobado nivel de evidencia, el proceso puede sintetizarse
de la siguiente manera:
Lo primero que aumenta
es el volumen corriente, que puede llegar a duplicarse. El organismo
evitaría usar volúmenes mayores ya que esto significaría
respirar en un nivel de mayor volumen pulmonar tiene menor distensibilidad,
o sea, se necesita generar mayores cambios de presión
por unidad de volumen lograda. Si es necesaria mayor ventilación
ésta se logra con un menor aumentando la frecuencia respiratoria.
Mejorada la oferta pulmonar de O2el
sistema circulatorio es el encargado de transportarla a los
músculos aumentando el volumen sistólico y frecuencia
cardíaca, lo que puede quintuplicar el volumen de sangre
que circula por minuto. Por vasoconstricción selectiva
la sangre se deriva de los demás órganos a los
músculos en acción que están vasoldilatados
, pudiendo cuadruplicar su perfusión, reclutando capilares
que en reposo están colapsados. La acidificación
y calor local, ligados a la contracción muscular,
también contribuyen a abrir capilares.
Similar efecto se observa en el pulmón
en el cual la mayor perfusión significa llevar sangre
a los vértices que en posición vertical en reposo
forman parte del espacio muerto alveolar. A pesar de esto, a
veces se puede producir un aumento de la diferencia alvéolo-arterial
porque la sangre venosa vuelve mas insaturada. que en reposo.
En ejercicio leve o moderado,
que significa un consumo de O2 alrededor del
50-60% del consumo máximo del individuo, los músculos
metabolizan principalmente la glucosa en forma aerobia, liberando
mucha energía (36 moléculas de ATP por molécula
de glucosa). Si ante un mayor o mas prolongado esfuerzo el aporte
O2 no sigue aumentando la metabolizacion de glucosa
se hace anaerobia liberándose mucho menos energía
(2 ATP por molécula de glucosa) y se forma ácido
láctico que, al ser tamponado por bicarbonato produce
más CO2. Este umbral anaeróbico depende
del nivel usual de actividad física de cada sujeto( en
un atleta puede ser de 80% de su consumo máximo)
y de la eficacia de su pulmón y sistema cardiovascular.
Durante el ejercicio, la PaCO2
no sólo no aumenta, sino que incluso puede disminuir
un poco si el ejercicio muy intenso. La PaO2
en general se eleva y el pH se mantiene casi constante hasta
el momento en que empieza a producirse ácido láctico.
En la génesis del aumento de la ventilación
se han identificado varios estímulos:
- Impulsos nerviosos que partirían
del encéfalo, al prepararse para el ejercicio.
-
Estímulos que se originan en las
extremidades durante el ejercicio y que son transmitidos
por vía nerviosa a los centros respiratorios. Los
movimientos pasivos de una extremidad en un animal anestesiado
provocan un aumento de la ventilación, fenómeno
que se mantiene aunque se liguen los vasos sanguíneos
de la extremidad, pero que cesa si se seccionan los nervios.
Este mecanismo y el anterior se consideran como anticipatorios,
ya que la ventilación aumenta más de lo que
exige la demanda metabólica con la consiguiente alcalosis
respiratoria.(Fase neurológica)
-
Aumento de la producción muscular
de CO2, que estimula
y aumenta la sensibilidad de los centros bulbares, con incremento
de la ventilación, manteniéndose así
niveles normales o levemente bajos de CO2
en la sangre arterial. (Fase metabólica)
-
Con la prolongación del ejercicio
, se suma la acumulación de metabolitos musculares,
como el ácido láctico, con la consiguiente
acidosis que exige hiperventilación compensatoria.
Esta situación es la principal responsable de la
mantención de una ventilación aumentada durante
el período de reposo después del ejercicio/Fase
compensatoria)
-
La disminución de contenido de
O2 arterial actuaría en ejercicios intensos
a nivel de sensores musculares que captan la baja de disponibilidad
local de oxígeno.
En sujetos normales cuando el ejercicio llega a a ser intolerable
aun es posible aumentar el volumen corriente y la frecuencia
respiratoria y, por lo tanto, la ventilación global .
En cambio los aspectos circulatorios como el débito sistólico,
frecuencia cardíaca y extracción de oxigeno en
los capilares coronarios han llegado o están muy cerca
de sus niveles máximos, por lo cual se constituyen en
el factor limitante del ejercicio. Diferente puede ser la situación
en casos de enfermedad pulmonar o de musculatura de las extremidades
alterada.
Terminado el ejercicio la hiperpnea persiste por algunos minutos
hasta que se cubre la deuda de O2 que significa el
metabolismo anaeróbico.
| Un
hecho de extrema importancia es que parte importante de
los mecanismos mencionados están mas desarrollados
en las personas con mayor actividad física y que
pueden recuperarse, en parte significativa , en los enfermos
mediante entrenamiento |
CONTROL DE LA VENTILACION DURANTE EL SUEÑO
Durante el sueño, la ventilación
disminuye más allá de lo esperable por la reducción
de las demandas metabólicas: cae 6-10% al pasar del
estado vigil a la etapa de sueño no-REM y un 16% al
pasar a la etapa REM (rápida eye movements),
con la consiguiente caída en la PaO2 y aumento
en la PaCO2.
Dos mecanismos contribuyen a la hipoventilación:
a) Aumento de la resistencia de la vía aérea
alta por hipotonía de los músculos
dilatadores de la faringe. En el individuo despierto, la vía
aérea central, que tendería a colapsarse por
efecto de la presión negativa inspiratoria intraluminal,
se mantiene abierta por estimulación de los músculos
dilatadores de la faringe y de la laringe. Durante el sueño,
la hipotonía normal de los músculos dilatadores
de la faringe provoca un leve aumento de la resistencia al
flujo de aire, lo que se traduce muchas veces en ronquidos
y, en hombres mayores de 40 años, en episodios cortos
y alejados de apnea obstructiva, tema que es abordado en el
Capítulo 57
b) Variación de la actividad de los centros respiratorio.
.
MEDICION DE LA ACTIVIDAD DE LOS CENTROS RESPIRATORIOS
Dentro del área de la investigación
y de la especialización la actividad de los centros
respiratorios se ha estudiado habitualmente midiendo el cambio
de la ventilación frente a un determinado estímulo,
como es la inhalación de mezclas con concentraciones
altas de CO2 o bajas
de O2. En condiciones de integridad
mecánica del aparato respiratorio, este método
resulta adecuado, pero no lo es cuando existen alteraciones
del efector toracopulmonar. En estas condiciones, una respuesta
ventilatoria disminuida puede deberse tanto a falla del efector
como a disminución de la actividad de los centros respiratorios.
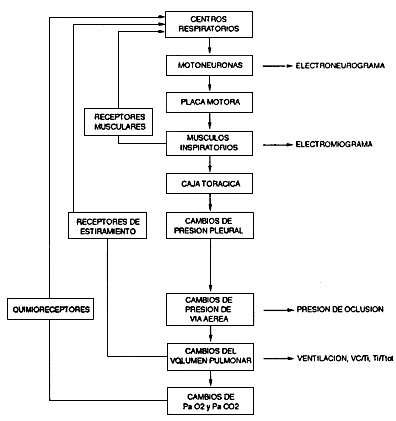
En la Figura 7-3 se muestra en forma esquemática la
serie de estructuras que intervienen en la generación,
transmisión y transformación del estímulo
en respuesta ventilatoria.
Figura 7-3. La ventilación
pulmonar es secundaria a la actividad automática
de los centros respiratorios, que se transmite a las motoneuronas
de los músculos inspiratorios; el impulso generado
por éstas se propaga por los nervios hacia las placas
motoras y a los músculos inspiratorios, cuya contracción
ensancha la caja torácica, lo que hace más
negativas las presiones pleural, alveolar y de vía
aérea, hecho que determina la entrada de aire al
pulmón. La actividad de los centros respiratorios
puede ser evaluada indirectamente mediante la medición
de cambios electroneurográficos, electromiográficos,
de la presión de vía aérea o del volumen
pulmonar, en relación con un estímulo químico
como cambios de PaO2
o PaCO2,
o mecánicos, como cambio en la resistencia de la
vía aérea o elastancia toracopulmonar.
En el esquema se destaca, además,
los exámenes que se pueden utilizar para medir la actividad
de cada nivel de la cadena. Se ha incluido también
los arcos reflejos que pueden modificar la actividad de los
centros respiratorios.
Los siguientes parámetros, propios
del área especializada, pueden utilizarse como índices
más directos de la actividad respiratoria central:
a) El electro-neurograma, que da información
experimental sobre la actividad de los centros respiratorios
por el registro de los potenciales de acción de un
nervio, por lo general del frénico.
b) La electromiografía de los músculos
inspiratorios, que tiene el inconveniente de proporcionar
información parcial porque es difícil estudiar
simultáneamente todos los músculos respiratorios.
c) La presión de oclusión (P
0.1), que es la presión generada por los músculos
inspiratorios durante el primer décimo de segundo de
una inspiración contra la vía aérea ocluida.
Para medirla, el sujeto respira a través de una válvula
cuya rama inspiratoria puede ocluirse sin que el sujeto lo
note. En este momento los músculos inspiratorios han
iniciado su contracción en forma proporcional al estímulo
central que han recibido: como se ha interrumpido el flujo
aéreo, no hay resistencias friccionales ni cambios
de volumen que interfieran en la presión desarrollada.
La presión de oclusión así medida es
muy reproducible y su valor promedio es de 1,6 ± 0,5
cmH2O.
Se ha demostrado que su medición es válida en
todo sujeto que sea capaz de generar una presión inspiratoria
máxima que supere los 20 cm H2O,
de modo que los trastornos musculares no la modifican, salvo
que sean muy avanzados. Tiene la ventaja sobre el neurograma
y electromiograma de entregar una información global,
ya que capta la activación simultánea de todos
los músculos inspiratorios.
|
|
|